
La mejora de la capacidad natural del tracto digestivo de los rumiantes para extraer nutrientes del alimento ha sido un objetivo importante para los nutricionistas durante más de medio siglo. No es demasiado sorprendente que el enfoque principal se haya centrado principalmente en los alimentos de menor digestibilidad. Sin embargo, el aumento de los precios de los piensos ha aumentado la necesidad de que los productores de ganado rumiante busquen nuevos programas biotecnológicos de alimentación que maximicen la utilización de los nutrientes de los piensos.
Un enfoque ha sido un intento de aumentar la capacidad digestiva del rumen estimulando un aumento en la microflora del rumen con la suplementación de cepas específicas de cultivos de levadura vivos viables que estimulan la producción de una importante bacteria digestiva de fibra, Ruminococcus albus (Dawson y Girard, 1997) .
Otro enfoque ha sido un intento de modificar los propios materiales de alimentación para hacerlos más fácilmente digeribles con enzimas. Recientemente, se han realizado intentos para definir las condiciones bajo las cuales las enzimas exógenas son más capaces de provocar una respuesta productiva positiva en rumiantes (Feng et al., 1996; Sanchez et al., 1996). En comparación con el uso en monogástricos, la investigación sobre la aplicación de la tecnología enzimática para su uso en la producción de rumiantes está en sus inicios. Este documento sugerirá cómo las enzimas pueden ser útiles en las dietas de los rumiantes y describirá los estudios de rendimiento y función del rumen en un nuevo suplemento de enzimas fibrolíticas, Fibrozyme.
Estabilidad de enzimas en el rumen
Al considerar la excepcional capacidad de digestión de fibra del rumen, es difícil ver cómo las enzimas fibrolíticas agregadas a los forrajes antes del consumo podrían mejorar aún más la utilización de los forrajes por parte de los rumiantes. Por otro lado, la digestión de forrajes en rumiantes es considerablemente menos que completa; y la tasa de digestión de la fibra tiene un gran impacto en el consumo, especialmente cuando se trata de forrajes tropicales. Las enzimas fibrolíticas suplementarias pueden actuar iniciando la degradación de los polisacáridos estructurales de la planta antes de la ingestión y la digestión ruminal, o complementando las enzimas fibrolíticas producidas por los microorganismos ruminales.
Hasta hace poco, se daba por sentado que las enzimas suplementarias que no se unían a su subestado objetivo serían rápidamente degradadas por las enzimas proteolíticas microbianas o del huésped en el tracto digestivo. Sin embargo, estas preocupaciones pueden resultar infundadas. Ahora se sabe que la extensa glicosilación de enzimas fúngicas las protege contra el ataque proteolítico en animales monogástricos (Chesson, 1993). Las mejoras recientes en la tecnología de enzimas han permitido la selección de mejores cepas de organismos y la identificación de enzimas con actividades principales y parciales más claramente definidas. Los productos de enzimas comerciales que se comercializan como aditivos para piensos se producen a partir de extractos de fermentación de origen bacteriano (Bacillus spp.) o fúngico (p. ej., Aspergillus, Trichoderma spp.).
Una excelente revisión de Beauchemin y Rode (1996) discutió los problemas de comparar y evaluar algunos de los estudios más antiguos con enzimas en raciones de rumiantes. Remitiríamos al lector a esta revisión para una mejor comprensión de las enzimas para rumiantes. Debe tenerse en cuenta que, en la actualidad, prácticamente toda nuestra experiencia con enzimas proviene de preparaciones bacterianas y fúngicas relativamente crudas. La preparación comercial normal de una enzima ‘celulasa’ puede contener una amplia variedad de actividades además de la celulasa. Algunas de estas actividades separadas pueden jugar un papel importante en la verdadera digestibilidad final de un sustrato.
Enzimas en acción
No está claro si el mayor beneficio de la aplicación de enzimas ocurre en el tratamiento previo a la alimentación o después de que el alimento ingresa al rumen. El beneficio también puede implicar cierto grado de protección de la enzima contra la descomposición microbiana del rumen. Las condiciones en el rumen pueden ser más constantes en cuanto a pH y temperatura y, por lo tanto, más ideales para la actividad enzimática. Sin embargo, también hay evidencia de que cuando las enzimas se agregan directamente al pasto y los ensilajes, tienen un beneficio definitivo para mejorar la digestibilidad de los forrajes. Por lo tanto, puede ser útil considerar la acción de las enzimas con respecto tanto al pretratamiento como a las aplicaciones ruminales o de alimentación directa.
Previo al tratamiento / Almacenamiento
El tratamiento del alimento o forraje mucho antes de la alimentación, como en las aplicaciones de inoculantes de ensilaje, permite el potencial para aumentar la actividad enzimática del rumen. Nakashima y Orskov (1989) informaron un aumento de hasta un 10% en la digestibilidad de la paja. Más recientemente, Beauchemin et al. (1995) que el tratamiento con enzimas de forrajes de alta calidad como la alfalfa y el pasto timothy durante el proceso de cubos podría aumentar significativamente el valor de los nutrientes para los novillos al aumentar la ganancia diaria promedio (ADG) en un 30 % y el consumo de materia seca (DMI) en un 10 %.
El tratamiento enzimático del alimento dentro de un corto período de tiempo (de unos pocos minutos a una hora) de la alimentación se describe como ‘alimentación directa’. A diferencia del pretratamiento enzimático, las enzimas de alimentación directa dependen de la respuesta dentro de un marco de tiempo más rápido y se espera que actúen durante el tránsito de la digesta. Este enfoque ofrece una mayor flexibilidad en el uso de enzimas en la granja y permitirá al agricultor maximizar el valor nutritivo de los cultivos forrajeros locales más baratos y, por lo tanto, reducir los costos de producción tanto como la fabricación de alimentos para aves o cerdos actualmente utiliza suplementos de enzimas.
Todavía existe una pregunta importante en cuanto a la posibilidad de que las enzimas sobrevivan en el rumen durante el tiempo suficiente y con una actividad suficiente para dar un efecto consistente en la digestibilidad de la fibra. Se informa que los niveles de enzimas fibrolíticas en el rumen son bajos (Vandevoorde y Verstaete, 1987). Fue informado por Kopecny et al. (1987) que la estabilidad de la celulasa en el rumen es baja. Sin embargo, Forwood et al. (1990) encontraron que cuando las enzimas líquidas se rociaron directamente sobre la festuca alta, se encontró un aumento del 5% en la digestibilidad de la materia seca in vitro. Lewis et al. (1996) también mostró un efecto positivo al agregar enzimas a las dietas basadas en forrajes antes de la alimentación. Este estudio, que investigó los efectos del tratamiento del forraje con enzimas 16 h antes de la alimentación, mostró un aumento en la producción de ácidos grasos volátiles (AGV) del 30 %; pero se observó poco efecto sobre la digestibilidad del forraje cuando la enzima se infundió directamente en el rumen. Por lo tanto, parecería que los medios para administrar la enzima en el rumen y el tiempo permitido entre el tratamiento y la alimentación pueden ser cruciales.
Dosis / respuesta
Estudios recientes han informado diferencias de rendimiento al complementar varios niveles de enzimas. Esto puede ayudar parcialmente a explicar algunas de las variaciones que se muestran en la literatura. Beauchemin et al. (1995) han demostrado que es posible sobretratar o subtratar con enzimas cuando se trata forraje para animales de carne. En un estudio reciente, Beauchemin y Rode (1996) señalaron que muchos experimentos que muestran los efectos de las enzimas han arrojado resultados tanto positivos como negativos. Estos mismos autores informaron en este estudio que cuando se rociaba una enzima sobre el forraje, encontraron un aumento en la GMD del 13% pero ninguna diferencia en el consumo de alimento. Una adición a este estudio, con una segunda preparación enzimática de actividades de xilanasa y celulasa, dio una respuesta de dosis positiva en ADG y tampoco mostró ningún efecto enzimático sobre el consumo de alimento (Tabla 1).
Nivel de dosis de enzima de investigación auricular por Sanchez et al. (1996) con vacas lecheras de alta producción alimentadas con heno de alfalfa utilizaron tres niveles de enzimas fibrolíticas (1.25, 2.5 y 5 l/tonelada de materia seca) como parte de una ración total mixta (TMR). Estos investigadores no mostraron una respuesta a la dosis cuando se administraron niveles más altos de enzimas. La respuesta enzimática positiva no fue lineal y fue más efectiva para el nivel intermedio de aplicación de enzimas en comparación con los animales de control que fueron alimentados con la ración no tratada (Tabla 2). Por el contrario, el DMI aumentó en todos los niveles de adición de enzimas. Los autores consideraron que la explicación de la menor producción de leche y el mayor consumo de alimento en el nivel más alto de aplicación de enzimas podría explicarse por un mayor puntaje de condición corporal que indica que las vacas estaban usando nutrientes para aumentar de peso en lugar de producir leche. Sin embargo, la diferencia en la producción de leche también podría deberse al emparejamiento desigual de vacas con las vacas en el grupo de tratamiento con alto contenido de enzimas que tienen un promedio de días en leche (DIM) más largo. Las diferencias en los patrones de ingesta esperados, así como en la producción de leche, explicarían en parte una dosis-respuesta inconsistente al tratamiento.
Tabla 1. Efecto de la dosis de enzima sobre la respuesta a las enzimas fibrolíticas en dietas de ensilado de maíz alimentadas al ganado.*
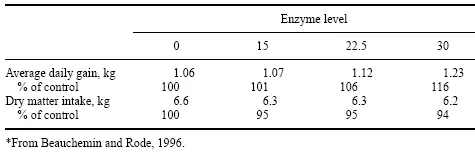
Tabla 2. Efectos en el rendimiento de vacas alimentadas con forraje tratado con tres niveles de enzimas.*
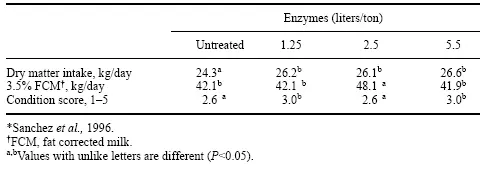
La diferencia en la producción de vacas lactantes emparejadas puede explicar gran parte de la respuesta inconsistente de la dosis de enzima para este ensayo, así como una gran parte de los datos informados en la literatura, especialmente cuando el número de cabezas de ganado es pequeño. Es por esta razón que queríamos observar un gran grupo de vacas de alta producción para poder comparar vacas con DIM similar e historial de producción para determinar si podíamos medir una diferencia de tratamiento efectiva en una nueva preparación de extracto de hongos para rumiantes ( fibrozima).
Efectos de la fibrozima en el desempeño de vacas lecheras
Raciones y prodedimientos para vacas de alta producción
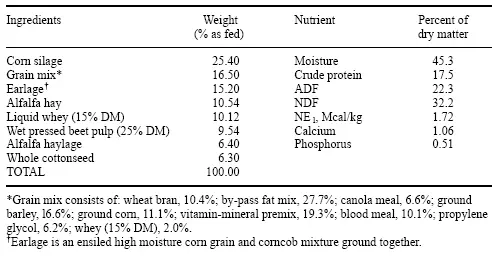
Se usaron más de 400 vacas Holstein de alta producción en una prueba de lactancia para evaluar los efectos de Fibrozyme. Las vacas fueron alimentadas con la misma TMR en seis corrales separados que contenían aproximadamente 100 animales por corral. Tres de los corrales se designaron como controles y tres corrales recibieron el suplemento de Fibrozyme a razón de 15 g/día. La composición de la TMR se da en la Tabla 3.
Tabla 3. Ingredientes y análisis de la ración.
Las vacas se emparejaron en función de la paridad, la producción, la etapa de lactancia y el uso de somatotropina bovina (bST). Se establecieron cuatro subgrupos separados de pares con respecto al tratamiento con bST. Las vacas que no recibieron bST durante todo el período de prueba se designaron como “sin bST” y las vacas que recibieron bST durante todo el período de prueba se denominaron “bST”. Las vacas originalmente asignadas a bST a las que se les quitó bST antes del final del experimento se denominaron ‘bST temprana’. De manera similar, las vacas asignadas al grupo ‘sin bST’ que posteriormente recibieron bST fueron designadas como el grupo ‘bST tardío’. Se hicieron ajustes por efecto de pluma, DIM y número de lactancia o antecedentes similares. Los datos se recopilaron del 5 de octubre al 7 de diciembre (63 días) y se analizaron estadísticamente para determinar las diferencias en la producción de leche, la grasa de la leche, la producción de leche corregida por la grasa (3,5 %) y la proteína de la leche.
Las muestras de producción de leche y componentes de la leche para los datos de vacas individuales se tomaron con muestreadores de leche Afikim para permitir la recolección de leche de principio a fin en cada unidad de ordeño. Se tomaron muestras de leche de vacas individuales en el ordeño de la mañana y de la tarde (6:30 a. m. y 2:30 p. m.) y se combinaron para analizar los componentes de la leche (grasa y proteína), así como el peso total de la leche. Las muestras de leche se enviaron al laboratorio DHIA en Ithaca, Nueva York, y el análisis estadístico se realizó en el Centro de Ciencias Lácteas de la Universidad de Florida.
Resultados
Fibrozyme tendió a aumentar la producción de leche mientras que bST aumentó la proteína de la leche (Tabla 4). Como las vacas no reciben bST hasta después de 100 a 120 días en la leche, lo que parece ser un efecto nulo de bST en la producción de leche no es el caso. Las vacas se emparejaron con efectos de tratamiento bST similares, de modo que se pudieran medir los rasgos de producción de leche. Las vacas que fueron emparejadas con el DIM promedio más corto (108) no recibieron bST, y las vacas que recibieron bST al final del período de prueba tuvieron el DIM promedio más largo (215). Históricamente, este rebaño ha informado un aumento del 15 % en la producción de leche para las vacas que reciben bST al final de la lactancia, cuando normalmente mostrarían una caída en la producción de leche.
Tabla 4. Efecto de Fibrozyme en vacas que recibieron diferentes tratamientos de bST.
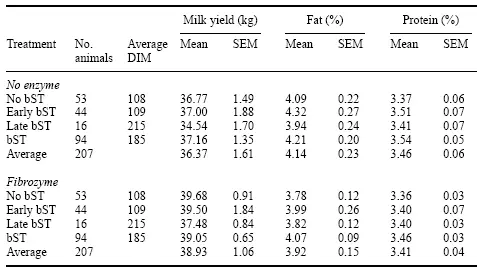
Fibrozyme tendió a aumentar la producción de leche independientemente de la asignación de bST (P < 0,08). La producción de leche más baja fue experimentada por el grupo control/bST tardío, aunque las diferencias no fueron significativas entre las asignaciones de bST. Los datos indicaron que bST aumentó el porcentaje de proteína (P<0,01), cuando se administró durante todo el ensayo. Fibrozyme aumentó la producción de leche durante el período de 63 días en un 7,04 % en comparación con el control.
Efectos de la fibrozima en la digestión ruminal de fibra
El tratamiento enzimático de los forrajes podría afectar beneficiosamente tanto la velocidad como la extensión de la digestión. Para examinar la dinámica de la digestión de forrajes tratados con enzimas, se realizó una serie de estudios in vitro en la Universidad de Kentucky.
Cuatro novillos fistulados fueron alimentados con una ración de 100% heno de pasto o 50% de concentrado suplementado con o sin Fibrozyme (15 g/día). El fluido ruminal de estos novillos se utilizó en los estudios in vitro que se describen a continuación. Cada animal recibió una dieta diferente durante cada uno de los cuatro períodos de muestreo. Los cuatro períodos de tratamiento consistieron en períodos de suplementación con enzimas de 8 días en los que los animales recibieron una dieta suplementada con enzimas o sin suplementos seguida de un período de descanso de 7 días en el que no se administraron suplementos. A mitad del ensayo se cambiaron los tratamientos dietéticos. El cambio de dieta estuvo acompañado de un período de adaptación de 14 días para que los animales se ajustaran a las nuevas dietas antes de iniciar la sucesión de la suplementación enzimática.
El fluido ruminal recolectado de cada animal una vez en cada período de suplementación se usó para evaluar la degradación in vitro de una dieta de referencia basada en festuca. El fluido ruminal se filtró a través de cuatro capas de gasa en dos botellas de 250 ml y se llevó de inmediato al laboratorio. Los frascos se introdujeron en una cámara anaeróbica (10% hidrógeno, 20% CO 2 , 70% nitrógeno) donde se mezcló el fluido ruminal con un volumen igual de saliva artificial de McDougall (McDougall, 1948). La mezcla se mezcló a alta velocidad durante 2 min y la suspensión celular resultante se usó como inóculo para los cultivos de frascos de suero. Todos los cultivos se prepararon en una fase gaseosa de CO2 en botellas de suero de 100 ml.
Se pesaron aproximadamente 0,5 g de una dieta molida de referencia en cada una de las 36 botellas. Los otros 36 no recibieron alimento y sirvieron como controles libres de sustrato. Una jeringa de 60 ml enjuagada con CO 2se utilizó para inocular 50 ml de suspensión celular en nueve frascos que contenían sustrato y nueve controles sin sustrato. Este procedimiento se repitió con las tres suspensiones celulares restantes hasta un total de 72 cultivos. Los cultivos se sellaron con un septo de goma y un sello de aluminio para mantener las condiciones anaeróbicas. Se incubaron en baño maría con agitación a 39°C. Se eliminaron tres cultivos replicados que contenían sustrato y tres controles sin sustrato de cada grupo de tratamiento después de 0, 12 y 24 h de incubación. Los pesos secos finales se corrigieron para el arrastre de materia seca de los inóculos de fluido ruminal utilizando los valores estimados de los controles sin sustrato en cada tiempo de incubación.
Producción de ácidos grasos volátiles
Las concentraciones de AGV se determinaron por cromatografía de gases. Se tomaron muestras del sobrenadante y se congelaron hasta su preparación. Se clarificaron con ácido metafosfórico al 25 % y se inyectó 1 ml en un cromatógrafo de gases Hewlett-Packard modelo 5890 serie II equipado con una columna de vidrio de 6 pies x 4 mm rellena con SP-1000 al 10 %/H 3 PO 4 al 1 % en 100/120 Chromosorb®WAW. La temperatura del horno se mantuvo constante a 135°C y el caudal fue de 32 ml/min. Los AGV producidos por cada cultivo se determinaron restando las concentraciones promedio de AGV a las 0 h de las concentraciones de AGV a las 12, 18 y 24 h. La utilización de hexosa se estimó estequiométricamente a partir de la producción de AGV calculando el balance de fermentación teórico.
Determinación de la desaparición de materia seca in vitro
La desaparición de materia seca se determinó para cada cultivo mediante una técnica de centrifugación. El material de los cultivos muestreados se lavó con 50 ml de agua destilada desionizada en botellas de centrifugación de 250 ml. Estos se centrifugaron a 4.000 xg durante 10 min y se descartó el sobrenadante.
El sedimento resultante se resuspendió en aproximadamente 5–10 ml de agua destilada desionizada y se transfirió a platos de aluminio previamente pesados. Estos se colocaron en el horno de secado a 98°C durante la noche. Los platos se equilibraron a temperatura ambiente en un desecador y se registraron los pesos secos. Para estimar el arrastre de materia seca de los inóculos de fluido ruminal, la cantidad de dieta de referencia añadida a cada botella de suero se restó de los pesos secos finales obtenidos a las 0 h. La diferencia de peso observada se atribuyó a la entrada de materia seca del contenido del rumen que se usó para inocular los cultivos de botellas de suero. Este valor se usó para corregir los pesos secos finales para el arrastre en todos los tiempos de incubación. La cantidad de materia seca digerida después de 12 y 24 h se estimó para cada cultivo de tratamiento restando los pesos secos finales corregidos de la cantidad de dieta de referencia pesada en cada cultivo. La desaparición de materia seca in vitro se expresó como un porcentaje y se calculó dividiendo la materia seca estimada digerida por la cantidad de dieta de referencia añadida a cada cultivo de botella de suero.
Resultados
Los efectos de la suplementación con Fibrozyme sobre la digestión de fibra in vitro de la dieta 100 % heno de pasto demostraron que a las 12 h, la desaparición de materia seca in vitro fue un 44 % mayor en los cultivos que recibieron Fibrozyme en comparación con el control sin suplemento (Tabla 5). La utilización estimada de hexosa y la producción neta de AGV en cultivos suplementados con Fibrozyme también tendieron a ser mayores que en cultivos sin suplemento después de 12 horas de incubación.
La adición de Fibrozyme a las dietas de novillos que consumieron una ración de concentrado al 50% con heno de pasto no alteró los patrones de desaparición de materia seca in vitro (Cuadro 6). Sin embargo, la producción de AGV y la utilización de carbohidratos fueron mayores en cultivos derivados de animales suplementados con Fibrozyme que en animales que recibieron raciones sin suplementos. Esto sugiere que Fibrozyme tuvo un impacto significativo en la digestibilidad de los componentes solubles que se encuentran en estas dietas de alta concentración.
Conclusiones
Para el éxito comercial, las enzimas para rumiantes tendrán que competir o complementar una variedad cada vez mayor de aditivos para piensos, así como bST. La tecnología tendrá que ofrecer un rendimiento constante como mínimo para el retorno en la granja. Estamos aprendiendo más sobre cómo los extractos de fermentación fúngica pueden modificar la producción de microorganismos del rumen y cómo podemos usar enzimas para producir cambios más positivos en los ácidos grasos de cadena corta y mejora de la producción de leche.
Tabla 5. Desaparición de materia seca in vitro, utilización estimada de hexosa y producción neta de AGV a partir de una ración de referencia a base de heno mediante cultivos discontinuos inoculados con contenido ruminal de animales alimentados con una dieta de heno 100 % de pasto con o sin Fibrozyme a las 12 y 24 h.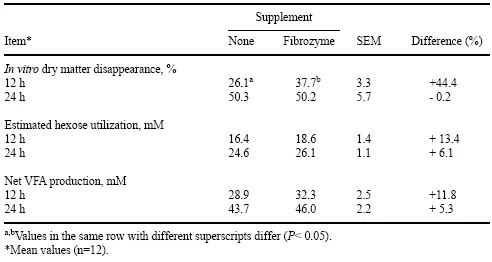
Tabla 6. Desaparición de materia seca in vitro, utilización de hexosas y producción neta de AGV a partir de una ración de referencia basada en heno mediante cultivos discontinuos inoculados con contenido ruminal de animales alimentados con una dieta concentrada al 50 % con o sin Fibrozyme a las 12 y 24 h.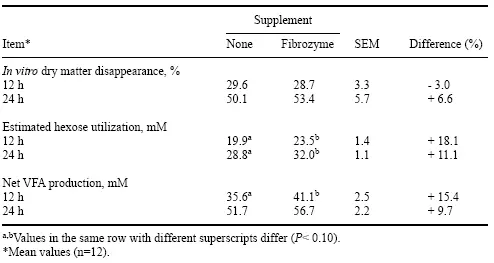
La respuesta a la suplementación con Fibrozyme en condiciones comerciales indicó que agregar enzimas para aumentar la digestión de la fibra tuvo un impacto positivo en la producción de leche de las vacas de alta producción. Los estudios in vitro indican que las adiciones de Fibrozyme a las dietas de los rumiantes mejoraron la digestión del material particulado y el metabolismo de los carbohidratos en una dieta de heno de festuca durante el corto período de incubación (primeras 12 h después de la adición), pero tuvieron pocos efectos consistentes a largo plazo sobre la digestión cuando se examinaron durante un período más largo (18-24 h después de la adición). El aumento en la tasa de digestión de 12 horas puede ayudar a explicar el aumento en la ingesta observado en respuesta a Fibrozyme en las pruebas de campo.
La capacidad del suplemento enzimático para mejorar los procesos digestivos parece depender de la población microbiana de origen y de la naturaleza de la dieta. Se observaron efectos máximos sobre la utilización del sustrato y la formación del producto final de la fermentación cuando se analizó la capacidad de las poblaciones microbianas obtenidas de animales alimentados con granos para utilizar carbohidratos en raciones basadas en forraje. La enzima también parece mejorar la capacidad de las poblaciones microbianas ruminales para degradar los carbohidratos solubles en animales alimentados con dietas que contienen concentrados y puede tener importantes aplicaciones especiales en sistemas donde se utilizan mezclas de concentrados y forrajes.
REFERENCIAS
Beauchemin, KA y LM Rode. 1996. Uso de enzimas alimenticias en la nutrición de rumiantes. Actas de la reunión anual de la Sociedad Canadiense de Ciencias Animales, Lethbridge, Alberta, del 7 al 11 de julio. págs. 103–130.
Beauchemin, KA, LM Rode y VJH Sewalt. 1995. Las enzimas fibrolíticas aumentan la digestibilidad de la fibra y la tasa de crecimiento de novillos alimentados con forrajes secos. Pueden. J. Anim. ciencia 75:641–644.
Chesson, A. 1993. Enzimas de alimentación. Tecnología científica de alimentación animal 45: 65–79.
Dawson, KA e ID Girard. 1997. Efectos estimulantes del cultivo de levadura en las bacterias del rumen. En: Biotecnología en la industria de piensos. Actas del 13º Simposio Anual (TP Lyons & KA Jacques, eds), Nottingham University Press, Loughborough, Leics, Reino Unido.
Feng, P., CW Hunt, GT Pritchard y WE Julien. 1996. Efectos de las preparaciones enzimáticas sobre las características digestivas in situ e in vitro del forraje de pasto maduro de estación fría en novillos de carne. J. Anim. ciencia 74:1349–1357.
Forwood, JR, DA Sleper y JA Henning. 1990. Efectos de la aplicación tópica de celulasa sobre la digestibilidad de la festuca alta. Agron. Revista 82: 909–913.
Kopecny, J., M. Marounek y K. Houb. 1987. Overenf vhodnosti aplikace celulaz Trichoderma viride do krmnych davek prezvykavcu [Prueba de la idoneidad de la adición de celulasas de Trichoderma viride a las raciones de pienso para rumiantes]. Zivocisna Vyroba 32:587–592.
Lewis, GE, CW Hunt, WK Sánchez, R. Treacher, GT Pritchard y P. Feng. 1996. Efecto de las enzimas fibrolíticas de alimentación directa sobre las características digestivas de una dieta a base de forraje alimentada a novillos de carne. J. Anim. ciencia 74:3020–3028.
McDougall, EI 1948. Estudios sobre saliva de rumiantes. I. La composición y producción de la saliva de las ovejas. Bioquímica J. 43:99.
Nakashima, Y. y ER Orskov. 1989. Degradación ruminal de paja – 7. Efectos del pretratamiento químico y la adición de ácido propiónico sobre las características de degradación de fracciones botánicas de paja de cebada tratada con una preparación de celulasa. Producción Animal 48:543–551.
Sánchez,WK, CW Hunt, MA Guy, GT Pritchard, B. Swanson, T.Warner y RJ Treacher. 1996. Efecto de las enzimas fibrolíticas en el desempeño de la lactancia de las vacas lecheras. Actas de la Asociación Americana de Ciencias Lácteas. Corvallis, Oregón. 14–17 de julio. (Póster).
Vandevoorde, L. y W. Verstaete. 1987. Fermentación anaeróbica en estado sólido con posible aplicación a la producción de celulasa. Aplicación J. Micro. Biotecnología. 26:479–484.
Autores: DEAN HOWES 1 , JUAN MARCELO TRICARICO 2 , KARL DAWSON 2 y PETER KARNEZOS 1
1 Alltech, Inc., Nicholasville, Kentucky, EE. UU
. 2 Departamento de Ciencias Animales, Universidad de Kentucky, Lexington, Kentucky, EE. UU.